Questo pezzo avrebbe potuto intitolarsi “Terroir come espressione fenotipica di Vitis vinifera L. a piede franco: superamento della discontinuità vascolare e dell’alterazione trascrittomica da portinnesto” ma per evitare a qualcuno possa venire in mente ci si voglia arrogare la pretesa di scrivere seriamente sul tema, quando in realtà dovrebbero essere ben più erudite penne a esprimersi, lo facciamo uguale, ma col titolo “Tutte le volte che si abusa del termne terroir” … l’ampelografia reale va a farsi friggere.
Il concetto moderno di terroir trova la sua validazione scientifica nell’espressione fenotipica di un genotipo in un determinato ambiente pedoclimatico. Storicamente, l’intera zonazione e la classificazione dei suoli viticoli d’eccellenza sono state codificate su piante a piede franco. Eppure, nel linguaggio comune del vino, la parola terroir è diventata un passe-partout: un termine inflazionato, svuotato del suo rigore e svenduto al marketing della suggestione, capace di offrire una qualità percepita del territorio priva di autenticità in valore assoluto.

Se vogliamo maneggiare questo concetto con cognizione di causa, dobbiamo spogliarlo della sua veste puramente geografica. Il terroir non è solo terra e clima: è un processo dinamico, uno spazio semantico dove convergono biologia, geologia e antropologia, senza mediazioni.
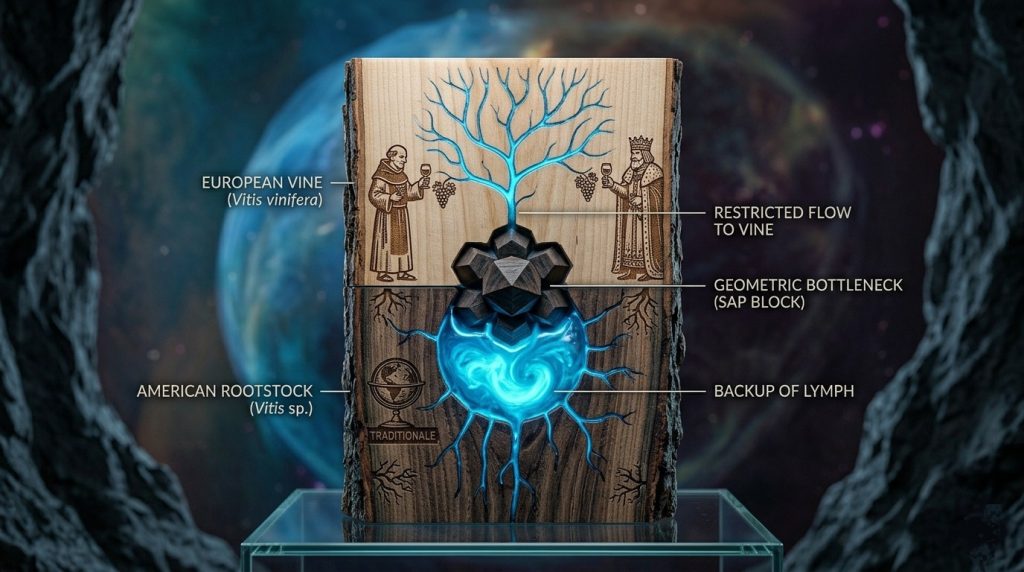
Tuttavia, l’enologia contemporanea ignora sistematicamente un presupposto fondamentale: affinché questo flusso relazionale si compia nella sua interezza, anche l’organismo trasmittente deve essere integro. L’avvento della fillossera e la conseguente introduzione del portinnesto americano hanno sì salvato la viticoltura, ma hanno al contempo inserito un elemento di alterazione permanente. Quella salvifica protesi sotterranea si configura oggi come un diaframma metafisico e biologico. Interrompendo la continuità originaria tra la Vitis vinifera e il suolo, l’innesto silenzia la frequenza più pura della terra, trasformando un dialogo millenario in un monologo squisitamente tecnologico, alterato e discontinuo a livello linfatico.
La comprensione profonda del terroir richiede un’archeologia del sapere viticolo. L’intera architettura dei grandi vini e le gerarchie dei cru storici sono nate prima della grande frattura fillosserica, fondandosi esclusivamente su piante a piede franco. Quando i monaci cistercensi mapparono i suoli della Borgogna, o quando l’editto del 1395 emanato da Filippo II l’Ardito bandì il Gamay definendolo un vitigno “molto cattivo e sleale” a favore del “buon vitigno” Pinot Noir, l’intento non era puramente agronomico, ma filosofico e organolettico allo stesso tempo. Si cercava l’espressione assoluta del luogo attraverso un organismo indiviso.
La stessa Classificazione del 1855 a Bordeaux ha cristallizzato il valore dei più grandi territori del mondo basandosi su vigne franche. Questo dimostra che l’idea stessa di terroir stampata nella memoria collettiva non è un’invenzione dell’enologia moderna, bensì il ricordo di un’armonia biologica perduta, dove la terra parlava direttamente all’acino senza intermediari genetici.
Sotto il profilo strettamente biologico, l’innesto rappresenta una barriera permanente nel sistema vascolare della pianta. La fusione tra il nesto e il portinnesto avviene attraverso un callo parenchimatico di cellule indifferenziate che genera una discontinuità nella connessione xilematica. Questo “collo di bottiglia” idraulico aumenta la resistenza interna e altera il flusso della linfa grezza, ostacolando il trasporto polare delle auxine e degli altri messaggeri chimici.

Dal punto di vista biochimico, l’apparato radicale della vite americana opera come un laboratorio endocrino estraneo: esso sintetizza citochine e acido abscissico secondo schemi genetici propri, inviando alla parte aerea segnali di maturazione e di gestione dello stress idrico parzialmente distorti. Il messaggio della terra viene così tradotto e modificato da un DNA estraneo prima di raggiungere il grappolo.
A livello nutrizionale, la letteratura scientifica evidenzia l’iper-efficienza del portinnesto nell’assorbire il potassio dal suolo. Tale eccesso di cationi si riversa nell’acino, dove l’acido tartarico viene salificato e precipita sotto forma di bitartrato. Il risultato biologico è un innalzamento sistematico del pH del mosto: il vino perde la sua naturale verticalità acida, costringendo la cantina a interventi correttivi massivi che simulano chimicamente una freschezza originaria che la pianta non è più in grado di esprimere.
Nelle istruzioni per l’uso del terroir, l’atto dell’assaggio diventa uno strumento di indagine filosofica e scientifica. Il vino da piante non innestate si distingue per tre precisi vettori sensoriali:
Omeostasi Acido-Base ed Efficacia Sensoriale: Il Vettore Idrogenionico
La conservazione di un pH naturalmente basso nel vitigno non innestato è l’effetto diretto di una regolazione cinetica controllata nell’assorbimento del catione. A livello organolettico, questa stabilità della concentrazione idrogenionica libera si traduce in una stimolazione lineare dei recettori cellulari e dei canali ionici (come i canali PKD2L1) deputati alla percezione della componente acida.
L’assenza di salificazione precoce dell’acido tartarico preserva la forza ionica intrinseca della soluzione idroalcolica. Dal punto di vista reologico e sensoriale, ciò determina una progressione gustativa continua e focalizzata, definibile come “tensione acida nativa”. Questo equilibrio dinamico sostiene la massa del vino senza generare la tipica risposta astringente o disorganizzata riscontrabile quando l’acidità totale viene corretta artificialmente mediante addizioni tardive in cantina.
Sincronia di Fase della Maturazione Polifenolica e Reologia dei Tannini
L’assenza della resistenza idraulica specifica indotta dal callo parenchimatico dell’innesto garantisce la continuità del flusso xilematico e l’integrità del potenziale idrico fogliare. Questa stabilità omeostatica previene i micro-arresti fotosintetici indotti da stress idrico artificiale, consentendo una sincronizzazione ottimale tra l’accumulo glucidico (maturazione tecnologica) e la polimerizzazione dei flavonoidi nei tessuti vacuolari del seme e della buccia (maturazione fenolica).
Ne consegue l’estrazione di proantocianidine ad alto grado di polimerizzazione media (mDP), strutturalmente stabilizzate e associate a frazioni polisaccaridiche endogene. Al palato, queste macromolecole manifestano una ridotta affinità di legame verso le proteine salivari (come la mucina e le PRP), attenuando il fenomeno dell’astringenza grezza e l’attivazione dei recettori dell’amaro (TAS2Rs). La frazione tannica risulta così perfettamente integrata, levigata e priva di monomeri fenolici liberi o composti derivanti da necrosi cellulari o arresti metabolici dell’acino.
Dinamiche Metabolomiche dello Scudo Antiossidante e Protezione del Corredo Volatile
L’integrità del sistema immunitario e radicale della vite a piede franco promuove una sovraespressione dei geni codificanti l’enzima stilbene sintasi, stimolando la biosintesi di composti della classe degli stilbenoidi, tra cui il trans-resveratrolo e i suoi oligomeri, le viniferine. Sotto il profilo chimico, queste molecole agiscono come sistemi redox endogeni ad altissima efficienza, capaci di intercettare i radicali liberi e le specie reattive dell’ossigeno prima che si inneschi la degradazione per via ossidativa.
Questa protezione endogena preserva l’integrità dei precursori aromatici primari, specificamente i monoterpeni liberi e i tioli varietali legati ai gruppi solfidrilici (come il 3-mercaptoesanolo o 3MH). La stabilità cinetica di questo scudo molecolare garantisce una netta purezza del profilo olfattivo e una resistenza intrinseca all’invecchiamento atipico.
Al contrario, la vite innestata, a causa dell’alterazione trascrizionale indotta dal portinnesto, presenta una minore dotazione endogena di questi antiossidanti naturali. Ciò rende la matrice instabile e costringe l’enologia interventista a ricorrere a correzioni esogene massive come, ad esempio, i tannini sacrificali e i derivati di lievito ricchi in glutatione, o a trattamenti fisici di rimozione del potassio, come le resine a scambio ionico, per simulare artificialmente la longevità nativa del piede franco.

Fisiologia molecolare della percezione: Meccanismi recettoriali del pH
La scomposizione sensoriale descritta non risponde a suggestioni estetiche, ma a precise cascate biochimiche che si attivano all’interno della cavità orale. L’assaggio di un vino da Vitis vinifera L. a piede franco stimola i recettori biologici in modo qualitativamente differente rispetto a un vino da pianta innestata, attivando una via di trasduzione del segnale verso il sistema nervoso centrale che il cervello traduce come un’esperienza di freschezza integrata.
Il pH naturalmente basso del piede franco attiva le cellule gustative di Tipo III presenti nei bottoni caliciformi della lingua. La compressione molecolare della freschezza avviene tramite una precisa sequenza elettrofisiologica:
Ingresso dei protoni: Gli ioni idrogeno liberi penetrano nel citoplasma delle cellule di Tipo III attraversando il canale selettivo per i protoni OTOP1 (Otopetrin-1).
Acidificazione citosolica e blocco del potassio: L’afflusso satura l’ambiente intracellulare, provocando la chiusura dei canali del potassio voltaggio-indipendenti (specificamente la famiglia Kir2.1), impedendo la naturale fuoriuscita di potassio dalla cellula.
Depolarizzazione di membrana: La ritenzione forzata dei cationi di potassio all’interno della cellula inverte il potenziale di membrana a riposo, innescando una depolarizzazione cellulare che apre i canali del calcio voltaggio-dipendenti.
Attivazione del complesso PKD2L1: Il canale PKD2L1 (Polycystic Kidney Disease 2-Like 1), un canale cationico non selettivo permeabile al calcio appartenente alla superfamiglia dei transitori di potenziale recettoriale (TRP), funge da modulatore termico, chimico e cinetico dell’impulso. Nelle cellule di Tipo III, il complesso formato da PKD2L1 invia un segnale polarizzato e ad alta frequenza al nervo della corda del timpano (chorda tympani).
Poiché il mosto a piede franco non ha subito la salificazione da potassio indotta dall’iper-efficienza radicale della vite americana, la concentrazione di protoni liberi stabili è massima. L’attivazione di OTOP1 e Kir2.1 è netta e priva di fluttuazioni ioniche. Il cervello riceve un segnale acido geometrico e pulito, non alterato da picchi salini che provocherebbero un’attivazione spuria dei recettori del salato o dell’amaro, e che si traduce nella percezione della “verticalità acida”.
Idrodinamica orale e affinità cinematica dei tannini: Il ruolo delle PRP salivari
L’assenza di micro-arresti fotosintetici nel piede franco, garantita da un flusso idraulico integro, permette la sintesi di proantocianidine ad alto Grado di Polimerizzazione Medio (mDP). La forma e le dimensioni di queste molecole polifenoliche determinano il comportamento reologico del vino a contatto con il fluido salivare.
Nelle maturazioni frammentate dei vigneti innestati, l’astringenza standard è definita dalla capacità dei tannini corti o monomerici di legarsi rapidamente alle proteine ricche di proline (PRP, Proline-Rich Proteins) e alle mucine che compongono il velo lubrificante della mucosa orale. Questo legame idrofobico causa la precipitazione delle proteine, aumentando l’attrito meccanico all’interno della cavità orale. Il cervello interpreta questo incremento d’attrito come una sensazione tattile rugosa, secca e allappante.
Al contrario, le macromolecole tanniche ad alto mDP, tipiche della vite non innestata, possiedono una struttura sterica estesa e flessibile. Quando entrano in contatto con le PRP della saliva, formano complessi colloidali stabili e solubili, anziché aggregati insolubili che precipitano immediatamente. La viscosità e la capacità lubrificante del velo salivare rimangono inalterate. I meccanocettori orali, come i corpuscoli di Meissner, non registrano un aumento della frizione o della rugosità. Al palato, la trama polifenolica viene percepita come vellutata e levigata, poiché il tessuto biologico non subisce lo shock meccanico da precipitazione proteica.
Oggi, per evitare derive ideologiche, la comunicazione del settore ha contrapposto il concetto di terroir assoluto a quello di terroir mediato o condizionato. Si è creata una scissione netta tra la realtà biologica della terra e la narrazione commerciale del vino, insomma, un tentativo di campo largo un tanto al chilo per giustificare l’assenza di coerenza semantica.
La stessa narrazione commerciale del vino potrebbe cominciare a parlare più spesso di acidificazione pre-fermentativa, resine a scambio ionico, utilizzo di ceppi di lievito acidificanti, tannini proantocianidinici ed ellagici, derivati di lievito ricchi in glutatione e di tutti quei processi che raddrizzano in cantina ciò che il portinnesto distorce in vigna. A lor piacendo s’intende. Preservare o reintrodurre il piede franco laddove le condizioni del suolo, tra cui sabbie, altitudini e matrici vulcaniche, lo consentono, non è un esercizio di romanticismo passatista. È una necessità scientifica e filosofica. Significa rifiutare l’omologazione del filtro tecnologico per ritrovare l’archetipo biologico del vino. Solo eliminando la protesi sotterranea possiamo restituire al terroir la sua funzione primaria: essere la voce nuda, pura e indomita della terra che si fa vino. È, in ultima analisi, l’unica via per cui il vino possa dirsi realmente, radicalmente, figlio della terra e della gente che quella terra non ha mai smesso di abitare e difendere.




 Il Capodanno del Mugnaio compie 10 anni: la celebrazione del territorio e della filiera agricola di Mulino Caputo
Il Capodanno del Mugnaio compie 10 anni: la celebrazione del territorio e della filiera agricola di Mulino Caputo  Tutte le volte che si abusa del termine “terroir”…
Tutte le volte che si abusa del termine “terroir”…  Toscana, il racconto delle “2 sponde d’Arno”
Toscana, il racconto delle “2 sponde d’Arno”  Cenando sotto un Cielo Diverso: a Villa Tony la XVIII edizione celebra “I Colori della Diversità”
Cenando sotto un Cielo Diverso: a Villa Tony la XVIII edizione celebra “I Colori della Diversità”  A Napoli Taverna La Riggiola presenta il suo menù estivo
A Napoli Taverna La Riggiola presenta il suo menù estivo